Syntéza amónnych solí v obličkách
V obličkách pôsobením aktívneho enzýmu glutaminázy dochádza k hydrolytickému štiepeniu glutamínu na kyselinu glutámovú a amoniak.
V obličkách dochádza pri vylučovaní protónov k súčasnej reabsorpcii iónov Na + a K +. Renálna glutamináza sa teda podieľa na regulácii acidobázickej rovnováhy.
Glutaminázová aktivita je závislá od pH. Pri acidóze sa zvyšuje aktivita enzýmu, čím sa zvyšuje vylučovanie protónov a znižuje sa stupeň acidózy.
U dospelých sa denne uvoľňuje 0,5-1,2 g amónnych solí. Tvoria 3,5 % vylúčeného dusíka. U detí relatívny podiel amónnych solí je vyšší ako u dospelých. U novorodencov je dusík amónnych solí až 8,5 %.
Biosyntéza močoviny a jej porušenie
Glutamín vstupuje do pečene, kde sa pôsobením enzýmu glutaminázy uvoľňuje amoniak, ktorý sa mení na močovinu. Aminokyselina ornitín sa podieľa na syntéze močoviny, preto sa syntéza močoviny nazýva ornitínový cyklus.

Hlavným miestom syntézy močoviny je pečeň.
Kľúčovými enzýmami pre biosyntézu močoviny sú karbamoylfosfátsyntetáza, ortinikarbamoyltransferáza a argináza. Zdrojmi dusíka v močovine sú amoniak a kyselina asparágová. Ornitínový cyklus úzko súvisí s Krebsovým cyklom. Kyselina fumarová z ornitínového cyklu „odchádza“ do Krebsovho cyklu. Cyklus trikarboxylových kyselín zase poskytuje ornitínovému cyklu molekuly ATP.



Vlastnosti vylučovania koncových dusíkatých produktov u detí
V detstve môže byť poruchy syntézy močoviny,čo vedie k zvýšeniu koncentrácie amoniaku v krvi a tkanivách. Prejavujú sa vracaním, kŕčmi po požití bielkovinových potravín. Príznaky intoxikácie sa zmierňujú frakčnou bielkovinovou výživou. Poruchy tvorby močoviny sú spojené s absenciou alebo nízkou aktivitou enzýmov zapojených do tohto procesu.
Afermentózy biosyntézy močoviny
Vo svaloch sa vyskytuje zvláštna väzba amoniaku - alanínový cyklus, znázornený na obrázku:

Otázka dokončená
metabolizmus dusíka
metabolizmus dusíka- súbor chemických premien, reakcií syntézy a rozkladu dusíkatých zlúčenín v organizme; zložka metabolizmu a energie. Pojem "metabolizmus dusíka" zahŕňa metabolizmus bielkovín(súbor chemických premien bielkovín a ich metabolických produktov v tele), ako aj výmena peptidov, aminokyseliny, nukleových kyselín, nukleotidy, dusíkaté bázy, aminocukry (pozri. uhľohydráty), obsahujúce dusík lipidov, vitamíny, hormóny a ďalšie zlúčeniny obsahujúce dusík.
Organizmus zvierat a ľudí prijíma stráviteľný dusík z potravy, v ktorej hlavným zdrojom dusíkatých zlúčenín sú bielkoviny živočíšneho a rastlinného pôvodu. Hlavným faktorom udržiavania dusíkovej bilancie – stavu AA, pri ktorom je množstvo vstupného a výstupného dusíka rovnaké – je dostatočný príjem bielkovín z potravy. V ZSSR denná sadzba bielkovín v strave dospelého človeka sa rovná 100 G, alebo 16 G proteínový dusík s energetickým výdajom 2500 kcal. Dusíková bilancia (rozdiel medzi množstvom dusíka, ktoré vstupuje do tela s jedlom a množstvom dusíka vylúčeného z tela močom, stolicou a potom) je indikátorom intenzity A. o. v tele. Hladovka alebo nedostatočná výživa dusíkom vedie k negatívnej dusíkovej bilancii, alebo k nedostatku dusíka, pri ktorom množstvo dusíka vylúčeného z tela prevyšuje množstvo dusíka vstupujúceho do tela s potravou. Pozitívna dusíková bilancia, pri ktorej množstvo dusíka zavedeného potravou prevyšuje množstvo dusíka vylúčeného z tela, sa pozoruje v období rastu tela, pri procesoch regenerácie tkanív atď. Stav A. o. do značnej miery závisí od kvality potravinového proteínu, ktorý je zase určený jeho zložením aminokyselín a predovšetkým prítomnosťou esenciálnych aminokyselín.
Všeobecne sa uznáva, že u ľudí a stavovcov A. o. začína trávením dusíkatých zlúčenín potravy v gastrointestinálnom trakte. V žalúdku dochádza k rozkladu bielkovín za účasti tráviacich proteolytických enzýmov. trypsín a gastrixín (pozri Proteolýza ) za vzniku polypeptidov, oligopeptidov a jednotlivých aminokyselín. Zo žalúdka vstupuje jedlo do dvanástnik a dolné časti tenkého čreva, kde sa peptidy podrobujú ďalšiemu štiepeniu katalyzovanému enzýmami pankreatickej šťavy trypsínom, chymotrypsínom a karboxypeptidázou a enzýmami črevnej šťavy aminopeptidázami a dipeptidázami (pozri nižšie). Enzýmy). Spolu s peptidmi. v tenké črevo rozkladajú komplexné bielkoviny (napr. nukleoproteíny) a nukleové kyseliny. Črevná mikroflóra tiež významne prispieva k rozkladu biopolymérov obsahujúcich dusík. Oligopeptidy, aminokyseliny, nukleotidy, nukleozidy atď. sa vstrebávajú v tenkom čreve, dostávajú sa do krvi a sú s ňou prenášané po celom tele. Proteíny telesných tkanív v procese neustálej obnovy tiež podliehajú proteolýze pôsobením tkanivových protses (peptidázy a katepsíny) a produkty rozkladu tkanivových bielkovín vstupujú do krvi. Aminokyseliny môžu byť použité na novú syntézu proteínov a iných zlúčenín (purínové a pyrimidínové bázy, nukleotidy, porfyríny atď.), na energiu (napríklad prostredníctvom začlenenia do cyklu trikarboxylových kyselín) alebo môžu byť podrobené ďalšej degradácii tvorba konečných produktov A. O., podliehajúcich vylučovaniu z tela.
Aminokyseliny, ktoré sú súčasťou potravinových bielkovín, sa používajú na syntézu bielkovín orgánov a tkanív tela. Podieľajú sa aj na tvorbe mnohých ďalších dôležitých biologických zlúčenín: purínové nukleotidy(glutamín, glycín, kyselina asparágová) a pyrimidínové nukleotidy (glutamín, kyselina asparágová), serotonín (tryptofán), melanín (fenylalpnín, tyrozín), histamín (histidín), adrenalín, norepinefrín, tyramín (tyrozín), polyamíny (arginín, metionín) , cholín (metionín), porfyríny (glycín), kreatín (glycín, arginín, metionín), koenzýmy, cukry a polysacharidy, lipidy atď. Najdôležitejšou chemickou reakciou pre organizmus, na ktorej sa podieľajú takmer všetky aminokyseliny, je transaminácia, ktorá spočíva v reverzibilnom enzymatickom prenose a-aminoskupiny aminokyselín na a-uhlíkový atóm ketokyselín alebo aldehydov. Transaminácia je základná reakcia v biosyntéze neesenciálnych aminokyselín v tele. Aktivita enzýmov, ktoré katalyzujú transaminačné reakcie, je aminotransferázy - má veľkú klinickú a diagnostickú hodnotu.
Degradácia aminokyselín môže prebiehať niekoľkými rôznymi cestami. Väčšina aminokyselín môže podstúpiť dekarboxyláciu za účasti dekarboxylázových enzýmov za vzniku primárnych amínov, ktoré sa potom môžu oxidovať v reakciách katalyzovaných monoaminooxidázou alebo diaminooxidázou. Pri oxidácii biogénnych amínov (histamín, serotonín, tyramín, kyselina g-aminomaslová) oxidázy tvoria aldehydy, ktoré podliehajú ďalším premenám, a amoniak, ktorého hlavnou cestou ďalšieho metabolizmu je tvorba močoviny.
Ďalšou hlavnou cestou degradácie aminokyselín je oxidačná deaminácia s tvorbou amoniaku a ketokyselín. Priama deaminácia L-aminokyselín u zvierat a ľudí prebieha extrémne pomaly, s výnimkou kyseliny glutámovej, ktorá je intenzívne deaminovaná za účasti špecifického enzýmu glutamátdehydrogenázy. Predbežná transaminácia takmer všetkých α-aminokyselín a ďalšia deaminácia vytvorenej kyseliny glutámovej na kyselinu α-ketoglutarovú a amoniak je hlavným mechanizmom deaminácie prirodzených aminokyselín.
Amoniak je produktom rôznych dráh degradácie aminokyselín, ktorý môže vzniknúť aj v dôsledku metabolizmu iných zlúčenín obsahujúcich dusík (napríklad pri deaminácii adenínu, ktorý je súčasťou nikotínamidadeníndinukleotidu – NAD). Hlavným spôsobom viazania a neutralizácie toxického amoniaku u ureotelických zvierat (zvieratá, u ktorých je konečným produktom A. o močovina) je takzvaný močovinový cyklus (synonymum: ornitínový cyklus, Krebsov-Henseleitov cyklus), ktorý prebieha v pečeni. . Ide o cyklickú sekvenciu enzymatických reakcií, v dôsledku ktorých sa z molekuly amoniaku alebo amidového dusíka glutamínu, aminoskupiny kyseliny asparágovej a oxidu uhličitého syntetizuje močovina. Pri dennom príjme 100 G proteín denné vylučovanie močoviny z tela je asi 30 G. U ľudí a vyšších živočíchov existuje ďalší spôsob neutralizácie amoniaku - syntéza amidov dikarboxylových kyselín asparagánu a glutamínu z príslušných aminokyselín. U urikotelických zvierat (plazy, vtáky) je konečný produkt A. o. je kyselina močová.
V dôsledku rozkladu nukleových kyselín a nukleoproteínov v gastrointestinálnom trakte vznikajú nukleotidy a nukleozidy. Oligo- a mononukleotidy za účasti rôznych enzýmov (esterázy, nukleotidázy, nukleozidázy, fosforylázy) sa potom premieňajú na voľné purínové a pyrimidínové bázy.
Ďalšia cesta degradácie purínových báz adenínu a guanínu spočíva v ich hydrolytickej deaminácii pod vplyvom enzýmov adenázy a guanázy za vzniku hypoxantínu (6-hydroxypurín) a xantínu (2,6-dioxipurínu), resp. sa potom premieňajú na kyselinu močovú v reakciách katalyzovaných xantínoxidázou. Kyselina močová je jedným z konečných produktov A. o. a konečný produkt metabolizmu purínov u ľudí – sa vylučuje z tela močom. Väčšina cicavcov má enzým urikázu, ktorý katalyzuje premenu kyseliny močovej na vylučovaný alantoín.
Degradácia pyrimidínových zásad (uracil, tymín) spočíva v ich redukcii za vzniku dihydroderivátov a následnej hydrolýze, v dôsledku ktorej z uracilu vzniká kyselina b-ureidopropiónová a z nej amoniak, oxid uhličitý a b-alanín, a kyselina b-aminoizomaslová z tymínu, kyselina uhličitá a amoniak. Oxid uhličitý a amoniak môžu byť ďalej zahrnuté do močoviny prostredníctvom močovinového cyklu a b-alanín sa podieľa na syntéze najdôležitejších biologicky aktívnych zlúčenín - dipeptidov obsahujúcich histidín karnozín (b-alanyl-L-histidín) a anserín (b -alanyl-N-metyl-L-histidín), ktorý sa nachádza v extraktívnych látkach kostrových svalov, ako aj pri syntéze kyseliny pantoténovej a koenzýmu A.
V jedinej výmene sú tak navzájom prepojené rôzne premeny najdôležitejších dusíkatých zlúčenín organizmu. Zložitý proces A. o. regulované na molekulárnej, bunkovej a tkanivovej úrovni. A. predpis o. v celom organizme je zameraná na prispôsobenie intenzity A. o. na meniace sa podmienky prostredia a vnútorného prostredia a uskutočňuje sa nervovým systémom priamo aj pôsobením na žľazy s vnútorným vylučovaním.
U zdravých dospelých jedincov je obsah dusíkatých zlúčenín v orgánoch, tkanivách a biologických tekutinách na relatívne konštantnej úrovni. Nadbytočný dusík z potravy sa vylučuje močom a stolicou a pri nedostatku dusíka v potrave je možné jeho potreby tela pokryť použitím zlúčenín dusíka v telesných tkanivách. Zároveň aj zloženie moču sa mení v závislosti od funkcií A. a dusíkovej bilancie. Normálne pri nezmenenej strave a relatívne stabilných podmienkach životné prostredie z tela sa vylučuje konštantné množstvo konečných produktov A. jazera a rozvoj patologických stavov vedie k jeho prudkej zmene. Významné zmeny vo vylučovaní dusíkatých zlúčenín v moči, predovšetkým vo vylučovaní močoviny, možno pozorovať aj pri absencii patológie v prípade významnej zmeny stravy (napríklad pri zmene množstva spotrebovaných bielkovín ), a koncentrácia zvyškového dusíka (pozri. Zvyškový dusík ) v krvi sa mierne mení.
Pri výskume And. je potrebné brať do úvahy kvantitatívne a kvalitatívne zloženie prijímanej potravy, kvantitatívne a kvalitatívne zloženie dusíkatých zlúčenín vylučovaných močom a stolicou a obsiahnutých v krvi. Pre výskum A. o. používať dusíkaté látky označené rádionuklidmi dusíka, fosforu, uhlíka, síry, vodíka, kyslíka a sledovať migráciu značky a jej začlenenie do zloženia konečných produktov A. o. Široko používané sú značené aminokyseliny, napríklad 15N-glycín, ktoré sa do tela dostávajú potravou alebo priamo do krvi. Významná časť označeného potravinového glycínového dusíka sa vylučuje močom ako močovina a druhá časť označenia vstupuje do tkanivových bielkovín a z tela sa vylučuje extrémne pomaly. Vykonávanie výskumu A. o. nevyhnutné pre diagnostiku mnohých patologických stavov a sledovanie účinnosti liečby, ako aj vývoj racionálnych diét, vr. liečivé (pozri Liečebná výživa ).
Patológia A. o. (až veľmi výrazný) spôsobuje nedostatok bielkovín. Môže to byť spôsobené všeobecnou podvýživou, dlhotrvajúcim nedostatkom bielkovín alebo esenciálnych aminokyselín v strave, nedostatkom sacharidov a tukov, ktoré poskytujú energiu pre procesy biosyntézy bielkovín v tele. Nedostatok bielkovín môže byť spôsobený prevahou procesov rozkladu bielkovín nad ich syntézou, a to nielen v dôsledku alimentárneho nedostatku bielkovín a iných základných živín, ale aj pri ťažkej svalovej práci, zraneniach, zápalových a dystrofické procesy, ischémia, infekcia, rozsiahle popáleniny, porucha trofickej funkcie nervový systém, nedostatok anabolických hormónov (rastový hormón, pohlavné hormóny, inzulín), nadmerná syntéza alebo nadmerný príjem steroidných hormónov zvonku a pod. Porušenie absorpcie proteínov v patológii gastrointestinálny trakt(zrýchlená evakuácia potravy zo žalúdka, hypo- a anacidné stavy, upchatie vývodu pankreasu, oslabenie sekrečnej funkcie a zvýšená pohyblivosť tenkého čreva pri enteritíde a enterokolitíde, porucha vstrebávania v tenkom čreve a pod.) viesť k nedostatku bielkovín. Nedostatok bielkovín vedie k poruche koordinácie A. o. a vyznačuje sa výraznou negatívnou dusíkovou bilanciou.
Sú známe prípady narušenia syntézy určitých proteínov (pozri. Imunopatológia, Fermentopatie), ako aj geneticky podmienená syntéza abnormálnych proteínov, napr hemoglobinopatie, mnohopočetný myelóm (pozri Paraproteinemické hemoblastózy ) a pod.
Patológia jazera A., ktorá spočíva v porušení metabolizmu aminokyselín, je často spojená s anomáliami v procese transaminácie: zníženie aktivity aminotransferáz počas hypo- alebo avitaminózy B 6, porušenie syntézy týchto enzýmov nedostatok ketokyselín na transamináciu v dôsledku inhibície cyklu trikarboxylových kyselín počas hypoxie a cukrovka atď. Zníženie intenzity transaminácie vedie k inhibícii deaminácie kyseliny glutámovej, a to zase k zvýšeniu podielu dusíka aminokyselín v zložení zvyškového dusíka v krvi (hyperaminoacidémia), všeobecnej hyperazotémii a aminoacidúrii. Hyperaminoacidémia, aminoacidúria a celková azotémia sú charakteristické pre mnohé typy A. patológie. Pri rozsiahlom poškodení pečene a iných stavoch spojených s masívnym rozkladom bielkovín v organizme sú procesy deaminácie aminokyselín a tvorby močoviny narušené tak, že sa zvyšuje koncentrácia zvyškového dusíka a obsah dusíka aminokyselín v ňom. na pozadí poklesu relatívneho obsahu močovinového dusíka v zvyškovom dusíku (tzv. produkčná azotémia).
Produkčná azotémia je zvyčajne sprevádzaná vylučovaním nadbytočných aminokyselín močom, keďže aj v prípade normálne fungovanie obličková filtrácia aminokyselín v glomerulách je intenzívnejšia ako ich reabsorpcia v tubuloch. Ochorenie obličiek, obštrukcia močové cesty zhoršená cirkulácia obličiek vedie k rozvoju retenčnej azotémie, sprevádzanej zvýšením koncentrácie zvyškového dusíka v krvi v dôsledku zvýšenia obsahu močoviny v krvi (pozri. zlyhanie obličiek ). Rozsiahle rany, ťažké popáleniny, infekcie, poškodenie tubulárnych kostí, miechy a mozgu, hypotyreóza, Itsenko-Cushingova choroba a mnohé iné vážnych chorôb sprevádzaná aminoacidúriou. Je tiež charakteristická pre patologické stavy, ktoré sa vyskytujú pri zhoršených procesoch reabsorpcie v obličkových tubuloch: Wilsonova-Konovalovova choroba (pozri. Hepatocerebrálna dystrofia ), Nephronophthisis Fanconi (pozri. Choroby podobné krivici ) a iné.Tieto ochorenia patria medzi početné geneticky podmienené poruchy A. o. Selektívne porušenie reabsorpcie cystínu a cystinúria s generalizovanou poruchou metabolizmu cystínu na pozadí všeobecnej aminoacidúrie sprevádza takzvanú cystinózu. Pri tomto ochorení sa cystínové kryštály ukladajú v bunkách retikuloendotelového systému. dedičné ochorenie fenylketonúria charakterizované porušením premeny fenylalanínu na tyrozín v dôsledku geneticky podmieneného nedostatku enzýmu fenylalanín - 4-hydroxylázy, ktorý spôsobuje hromadenie nepremeneného fenylalanínu a jeho metabolických produktov - kyseliny fenylpyrohroznovej a fenyloctovej v krvi a moči. Porušenie transformácií týchto zlúčenín je tiež charakteristické pre vírusovú hepatitídu.
Tyrozinémia, tyrozinúria a tyrozinóza sú zaznamenané pri leukémii, difúzne ochorenia spojivové tkanivo(kolagenózy) a iné patologické stavy. Vyvíjajú sa v dôsledku narušenej transaminácie tyrozínu. Základom alkaptonúrie je vrodená anomália oxidačných premien tyrozínu, pri ktorej sa v moči hromadí nepremenený metabolit tejto aminokyseliny, kyselina homogentisová. Poruchy metabolizmu pigmentov pri hypokorticizme (pozri. nadobličky ) sú spojené s inhibíciou premeny tyrozínu na melanín v dôsledku inhibície enzýmu tyrozinázy (úplná strata syntézy tohto pigmentu je typická pre vrodená anomália pigmentácia – albinizmus).
Chronická hepatitída, diabetes mellitus, akútna leukémia, chronická myelo- a lymfocytová leukémia, lymfogranulomatóza, reumatizmus a sklerodermia, metabolizmus tryptofánu je narušený a v krvi sa hromadia jeho metabolity kyseliny 3-hydroxykynurenín, xanturénová a 3-hydroxyantranilová, ktoré majú toxické vlastnosti. K A. patológii o. zahŕňajú aj stavy spojené s porušením vylučovania kreatinínu obličkami a jeho akumuláciou v krvi. Zvýšené vylučovanie kreatinínu sprevádza hyperfunkciu štítna žľaza a zníženie vylučovania kreatinínu so zvýšeným vylučovaním kreatínu je hypotyreóza.
S masívnym kolapsom bunkových štruktúr(hladovanie, ťažká svalová práca, infekcie atď.) Zaznamenajte patologické zvýšenie koncentrácie zvyškového dusíka v dôsledku zvýšenia relatívneho obsahu dusíka kyseliny močovej v ňom (normálne koncentrácia kyseliny močovej v krvi nepresahuje - 0,4 mmol/l).
V starobe sa intenzita a objem syntézy bielkovín znižuje v dôsledku priamej inhibície biosyntetickej funkcie tela a oslabenia jeho schopnosti absorbovať aminokyseliny z potravy; vzniká negatívna dusíková bilancia. Poruchy metabolizmu purínov u starších ľudí vedú k hromadeniu a ukladaniu solí kyseliny močovej – urátov vo svaloch, kĺboch a chrupavkách. Korekcia porúch A. v starobe sa môže vykonávať prostredníctvom špeciálnych diét s obsahom vysoko kvalitných živočíšnych bielkovín, vitamínov a stopových prvkov s obmedzeným obsahom purínov.
Metabolizmus dusíka u detí sa vyznačuje množstvom znakov, najmä pozitívnou dusíkovou bilanciou nevyhnutná podmienka rast. Intenzita procesov A. o. počas rastu dieťaťa prechádza zmenami, najmä u novorodencov a malých detí. Počas prvých 3 dní života je dusíková bilancia negatívna, čo sa vysvetľuje nedostatočným príjmom bielkovín z potravy. V tomto období sa zisťuje prechodné zvýšenie koncentrácie zvyškového dusíka v krvi (tzv. fyziologická azotémia), niekedy až 70 mmol/l; do konca 2. týždňa.
života, koncentrácia zvyškového dusíka klesá na úroveň zaznamenanú u dospelých. Množstvo dusíka vylučovaného obličkami sa zvyšuje počas prvých 3 dní života, potom klesá a od 2. týždňa sa opäť začína zvyšovať. života súbežne so zvyšujúcim sa množstvom potravy.
Najvyššia stráviteľnosť dusíka v tele dieťaťa sa pozoruje u detí v prvých mesiacoch života. Dusíková bilancia sa v prvých 3-6 mesiacoch výrazne približuje k rovnováhe. život, hoci zostáva pozitívny. Intenzita metabolizmu bielkovín u detí je pomerne vysoká – u detí 1. roku života asi 0,9 G proteín za 1 kg telesná hmotnosť za deň, za 1-3 roky - 0,8 g/kg/ dní, u detí predškolského a školského veku - 0,7 g/kg/ deň
Priemerná hodnota potreby esenciálnych aminokyselín je podľa FAO WHO (1985) u detí 6-krát vyššia ako u dospelých (esenciálna aminokyselina pre deti do 3 mesiacov je cystín a do 5 rokov - a histidín). U detí prebiehajú procesy transaminácie aminokyselín aktívnejšie ako u dospelých. V prvých dňoch života novorodencov sa však v dôsledku relatívne nízkej aktivity niektorých enzýmov v dôsledku funkčnej nezrelosti obličiek zaznamenáva hyperaminoacidémia a fyziologická aminoacidúria. U predčasne narodených detí navyše existuje aminoacidúria typu preťaženia, tk. obsah voľných aminokyselín v plazme ich krvi je vyšší ako u donosených detí. Aminokyselinový dusík tvorí v prvom týždni života 3 – 4 % celkového dusíka v moči (podľa niektorých zdrojov až 10 %) a až do konca 1. roku života jeho relatívny obsah klesá na 1 %. U detí 1. roku života je vylučovanie aminokyselín na 1 kg telesná hmotnosť dosahuje hodnoty ich vylučovania u dospelého človeka, vylučovanie dusíka aminokyselín, u novorodencov 10 mg/kg telesná hmotnosť, v 2. roku života zriedka presahuje 2 mg/kg telesná hmotnosť. V moči novorodencov je zvýšený obsah taurínu, treonínu, serínu, glycínu, alanínu, cystínu, leucínu, tyrozínu, fenylalanínu a lyzínu (v porovnaní s močom dospelého človeka). V prvých mesiacoch života sa v moči dieťaťa nachádza aj etanolamín a homocitrulín. V moči detí 1. roku života prevládajú aminokyseliny prolín a [hydro]oxyprolín.
Štúdie najdôležitejších dusíkatých zložiek moču u detí ukázali, že pomer kyseliny močovej, močoviny a amoniaku sa počas rastu výrazne mení. Áno, prvé 3 mesiace. života sa vyznačujú najnižším obsahom močoviny v moči (2-3x menej ako u dospelých) a najvyšším vylučovaním kyseliny močovej. Deti v prvých troch mesiacoch života vylučujú 28.3 mg/kg telesná hmotnosť kyseliny močovej a dospelí - 8,7 mg/kg. Pomerne vysoké vylučovanie kyseliny močovej u detí počas prvých mesiacov života niekedy prispieva k rozvoju infarktu obličiek z kyseliny močovej. Množstvo močoviny v moči sa u detí vo veku 3 až 6 mesiacov zvyšuje a obsah kyseliny močovej v tomto období klesá. Obsah amoniaku v moči detí v prvých dňoch života je malý, potom však prudko stúpa a zostáva na vysokej úrovni počas celého 1. roku života.
charakteristický znak A. o. u detí je fyziologická kreatinuria. Kreatín sa nachádza v plodovej vode; v moči sa stanovuje v množstvách prevyšujúcich obsah kreatínu v moči dospelých, od novorodeneckého obdobia až po obdobie puberty. Denné vylučovanie kreatinínu (dehydroxylovaného kreatínu) sa zvyšuje s vekom, pričom súčasne so zvyšovaním telesnej hmotnosti dieťaťa klesá relatívny obsah kreatinínového dusíka v moči. Množstvo kreatinínu vylúčeného močom za deň u donosených novorodencov je 10-13 mg/kg u predčasne narodených detí 3 mg/kg, u dospelých nepresahuje 30 mg/kg.
Pri identifikácii v rodine vrodených porúch A. potrebu
Krátka otázka
Izolácia konečných produktov metabolizmu dusíka
Kyselina močová je jedným z najdôležitejších konečných produktov metabolizmu dusíka u ľudí. Bežne je jeho koncentrácia v krvnom sére u mužov 0,27-0,48 mmol-l1, u žien 0,18-0,38 mmol-l-1; denné vylučovanie močom sa pohybuje od 2,3 do 4,5 mmol (400-750 mg). Ľudia vylučujú kyselinu močovú a mnohé cicavce majú enzým urikázu, ktorý oxiduje kyselinu močovú na alantoín. V tele zdravého človeka za deň sa tvorba a vylučovanie kyseliny močovej pohybuje od 500 do 700 mg. Väčšina kyseliny močovej (až 80 %) vzniká v dôsledku metabolizmu endogénnych nukleových kyselín, len asi 20 % je spojených s purínmi z potravy. Obličky vylúčia denne asi 500 mg kyseliny močovej, 200 mg sa odstráni cez gastrointestinálny trakt.
Funkčná proteinúria. Funkčná proteinúria, ktorej presné procesy výskytu neboli stanovené, zahŕňa tie, ktoré súvisia s vertikálnou polohou tela, idiopatická nestála, vylučovanie bielkovín v moči kmeňa, horúčkovitý výskyt bielkovín v moči a vylučovanie bielkovín v moči pri obezite.
Ortostatická proteinúria je charakterizovaná objavením sa polypeptidu v moči počas dlhšej nečinnosti alebo stimulácie, s jeho rýchlym vymiznutím, keď sa držanie tela zmení na kolmé. Výskyt proteínu v moči vo väčšine prípadov nepresahuje jeden g / deň, je glomerulárny a neselektívny, postup jeho výskytu je nejasný. Častejšie sa zaznamenáva v dospievaní, u polovice pacientov sa po chvíli vylieči. Mechanizmus tvorby je pravdepodobne spojený s abnormálne zvýšenou reakciou renálneho obehu na zmenu umiestnenia trupu.
Definícia ortostatickej proteinúrie je stanovená kombináciou nasledujúcich podmienok:
Vek pacientov 13-20 rokov;
Uzavretý typ proteínu v moči, absencia iných príznakov poškodenia obličiek (reštrukturalizácia močového sedimentu, zvýšenie tlaku, ktorým krv v tepne pôsobí na jej stenu, zmeny v cievach vnútorného povrchu očná buľva);
Len ortostatický priebeh výskytu bielkovín v moči, keď v štúdiách moču odobratého po tom, čo bol subjekt v polohe na chrbte (vrátane nasledujúceho rána pred vstávaním z postele), nie je žiadny proteín.
Na potvrdenie tejto diagnózy je potrebné vykonať test vo vzpriamenej polohe. Na tento účel sa moč odoberá ráno pred vstávaním z postele, potom po určitom čase v kolmej polohe (pohybom s palicou za chrbtom chrbtica). Diagnostika dáva ešte presnejšie výsledky, keď sa ranná (nočná) časť moču zlúči (pretože pri vesica urinaria je možný zvyškový moč) a počiatočná časť sa vyberie po krátkej prítomnosti pacienta v polohe na chrbte.
V mladom veku je zasa možný primárny intermitentný výskyt bielkovín v moči, ktorý sa u zdravých jedincov zistí pri lekárskej prehliadke a vymizne pri nasledujúcich vyšetreniach moču.
Tenzia proteinúria sa zistí u dvadsiatich percent zdravých jedincov (aj športovcov) po silnej fyzická aktivita. Proteín sa zisťuje v počiatočnej pripravenej časti moču. vylučovanie bielkovín v moči v prírode spojenej s patológiou tubulov. Predpokladá sa, že algoritmus na objavenie sa proteínu v moči je kombinovaný s rekombináciou prietoku krvi a relatívnou ischémiou proximálnych častí nefrónu.
Horúčkový výskyt bielkovín v moči sa vyskytuje v ťažkých horúčavách, najmä u detí a senilných ľudí. Horúčkové vylučovanie bielkovín močom má prevažne glomerulárny priebeh. Procesy tohto typu proteinúrie sú málo študované, možný význam sa zvyšuje glomerulárnej filtrácie spolu s krátkodobou porážkou glomerulárneho filtra ochrannými komplexmi.
Izolácia bielkovín v moči pri patologickej nadváhe. Vylučovanie bielkovín močom sa často pozoruje pri abnormálnom ukladaní tuku v tele. (telesná hmotnosť nad 115 kilogramov). Podľa J.P.Domfelda (1989) medzi tisíckou pacientov s patologickým ukladaním tuku v tele. 420 bolo diagnostikovaných s vylučovaním bielkovín močom bez degenerácie močových kalov; ukázané sú aj precedensy nefrotického syndrómu. Predpokladá sa, že hlavnou príčinou vzniku takejto proteinúrie je narušenie krvného obehu akumulácia vysoko fenestrovaných kapilár (zvýšený tlak v skupine kapilár obličkového telieska, zvýšená rýchlosť filtrácia) spojené s nárastom patologickej nadváhy. koncentrácia polypeptidového hormónu produkovaného obličkami v dôsledku poklesu v krvný tlak a hypertenzia, ktorá počas hladovania klesá. Pri úbytku hmotnosti, ako aj počas liečby ACE inhibítormi sa môže znížiť vylučovanie bielkovín močom, ako aj stratiť.
Okrem toho, proteinúria môže mať extrarenálny začiatok. V prítomnosti leukocytov v teste moču a najmä pri výskyte krvi v moči môže byť potvrdzujúcou reakciou na polypeptid výsledkom rozpadu krviniek počas dlhšieho státia v moči, v tejto situácii sa môže objaviť bielkovina v moči. moč presahujúci 0,3 gramu/deň sa zdá byť abnormálny. Sedimentárne polypeptidové testy môžu poskytnúť falošne pozitívne výsledky v prítomnosti kontrastných látok obsahujúcich jód v moči, značného počtu podobných prípravkov Penicillinum, ako aj liečivej látky zo skupiny polosyntetických beta-laktámových antibiotík, metabolických produktov sulfanilamidové prípravky.
Podobné informácie.
"Zbytkový dusík" Kjeldahlova metóda: v sére
krvi bola určená celk
vypočítalo sa množstvo dusíka a obsah bielkovín (v
bielkoviny ~ 16 % dusíka).
Po
depozícia
veverička
reakcia s TCA v sére
dusík zostal v
zloženie s nízkou molekulovou hmotnosťou
– „zvyškový (nebielkovinový)
dusík" (~ 0,5 % celkového N,
približne 0,4 g/l).
Močovinový dusík
50 %
Aminokyseliny dusíka
až 25%
Kyselina močová
4%
Kreatín a kreatinín
5-7 %
indický
0,5 %
Amoniak
<2%
Peptidy, nukleotidy,
bilirubín atď.
~13 %
Konečné produkty metabolizmu dusíka
VeveričkyAMC
Amines
Nukleové kyseliny
Pyrimidíny
AMONIAK
Kreatín
puríny
Močové
to-ta
Kreatinín
dočasný detox
amidy aminokyselín
s močom
UREA
Amoniak a spôsoby jeho neutralizácie
Deamináciaaminokyseliny
Deaminácia
biogénne amíny
Rozpad pyrimidínových báz NK
NH3
Syntéza AK,
pyrimidíny,
aminocukry…
Amidy dikarboxylových kyselín
aminokyseliny (hln, asn)
Vzdelávanie
a vylučovanie
amónny
soli
~ 0,5 g/deň
Syntéza
močovina
25-30 g/deň
Tvorba amidov dikarboxylových aminokyselín
Dočasná neutralizácia NH3 pre transport vorgány, kde je jeho konečná likvidácia a
vylučovanie.
Vyskytuje sa vo všetkých orgánoch a tkanivách.
vylučovanie amoniaku v obličkách
NH3 v moči sa viaže na protóny a vytvára katiónamoniak:
NH3 + H+ ↔ NH4+
NH4+ podporuje vylučovanie iónov H+ (t.j. kyselín).
Odstránenie amónnych solí (fosfáty, acetáty, ...)
znižuje straty Na a iných katiónov.
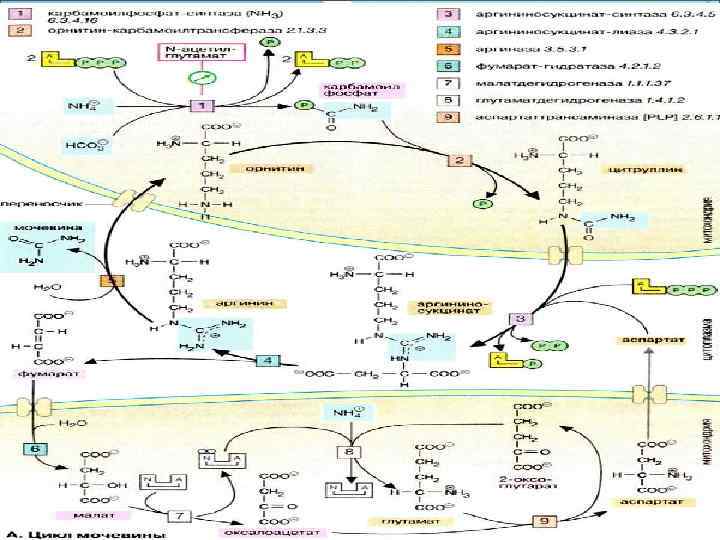
Syntéza močoviny
NH3Syntéza močoviny
ATP
CO2
NH3
Karbamoylfosfát
CITRULLÍN
ASPARTAT
Kyselina arginínjantárová
ORNITIN
Oxalacetát
Malat
ARGINÍN
NH2
OD
NH2
O
UREA
ATP
FUMARATE
močovina v krvi
Rýchlosťvzdelanie
v pečeni
UREA
KRV
Rýchlosť
odstránenie
obličky
Referenčné limity - 2,2 - 8,3 mmol / l. Denné výkyvy
hladina močoviny v krvi dosahuje 50 % (maximálne večer).
Schopnosť syntetizovať močovinu je zachovaná pri
poškodenie 85 % pečeňového tkaniva. Syntéza
močovina
zlomené len pri veľmi ťažkom poškodení pečene
(akútna nekróza, hepatálna kóma, cirhóza, otrava
fosfor a arzén): potom v krvnom sére
hromadí sa amoniak a hladina močoviny klesá.
stanovenie močoviny
Zvýšenie hladiny močoviny v krvi:relatívna (dehydratácia) + absolútna (AZOTÉMIA).
Azothemia
VÝROBA
ZACHOVANIE
zväčšené
vzdelanie
znížený
vylučovanie močom
RENÁLNA
EXTRARENAL
spojené s patológiou
obličky
spojené s extrarenálnymi faktormi
Produkčná azotémia
1.2.
Diéta bohatá na bielkoviny
Zvýšený katabolizmus bielkovín - s kachexiou, leukémiou,
rozsiahle rany, infekcie a
zápalové
choroby s vysokou horúčkou, malígne
nádorov
liečbe
glukokortikoidy,
intenzívne
svalová práca, AKÉKOĽVEK AKÚTNE FÁZOVÉ REAKCIE...
Retenčná azotémia
Ochorenie obličiek (GFR< 10 мл/мин) - гломерулонефриты,
pyelonefritída, renálna tuberkulóza, renálna amyloidóza…
Pri akútnom zlyhaní obličiek sa zvyšuje hladina močoviny v krvi až na 16 mmol / l =
stredne závažná porucha funkcie obličiek do 33 mmol / l -
ťažké, nad 50 mmol/l – veľmi závažné (nežiaduce
predpoveď).
Rastúce hladiny močoviny nie sú skorým príznakom dysfunkcie
obličky.
Retenčná azotémia
Extrarenálne retenčné azotémiehemodynamika a zníženie GFR:
-
pri
priestupkov
kardiovaskulárna dekompenzácia,
dehydratácia organizmu (nekontrolovateľné vracanie, obštrukcia
črevá, stenóza pyloru, profúzna hnačka, krvácanie,
popáleniny...)
traumatický šok, cukrovka, Addisonova choroba atď.
Azotémia je zvyčajne nízka (hladina močoviny< 13 ммоль/л).
Znížená koncentrácia močoviny
Nemá žiadnu diagnostickú hodnotu. Možno pozorovať:Pri hyperhydratácii (v / pri zavádzaní roztokov glukózy atď.)
Počas tehotenstva (často pod 3,33 mmol / l).
So zvýšenou diurézou (diuretikum).
Pri hladovaní a zníženom katabolizme bielkovín.
S poškodením svalov (myozitída, myopatia).
Metódy stanovenia močoviny
Kolorimetrické, ureáza (fenol-chlórnan, salicylát-chlórnan, glutamátdehydrogenáza).
Materiálom pre štúdiu je sérum alebo krvná plazma.
Hladina močoviny je stabilná až 24 hodín pri izbovej teplote,
niekoľko dní pri 4-6°C a až 2-3 mesiace pri zmrazení.
Rušenie
Koncentrácia močoviny v sére klesá počas hladovania,nízkokalorická strava, fajčenie, pitie veľkého množstva
voda…
Koncentrácia močoviny sa s vekom mierne zvyšuje. U mužov
je o niečo vyššia ako u žien.
Počas tehotenstva sa koncentrácia močoviny znižuje, v menopauze
- zvyšuje.
Diéta s vysokým obsahom bielkovín, cvičenie
spôsobiť zvýšenie koncentrácie močoviny v krvnom sére.
Nadhodnotenie výsledkov: acetón, bilirubín, hemolýza, lipémia,
oxaláty (prímes amónnych iónov), kyselina močová
+ nefrotoxické lieky, insekticídy.
Nedostatočné hlásenie: acidóza (metóda diacetylmonooxímu).
Kreatín a kreatinín
Kreatín je dôležitá zložkametabolizmus dusíka v tele.
Kreatín fosfát
zapojené
v
zásobovanie energiou
škrty
sval,
aktívny
dopravy
ióny v nervovom tkanive atď.
Syntéza kreatínu
Kreatín sa syntetizuje v obličkách a pankreasearginín, glycín a metionín a potom sa dostáva do krvného obehu
v kostrových a srdcových svaloch, mozgu, nervovom tkanive.
Obsah kreatínu v orgánoch
Obličky, pečeň,
LJ
Mozog
Hladké svaly
srdcový sval
Kostrové svaly
0
100
200
300
400
500
600
mg/100
Malé množstvo kreatínu (0,05-0,25 g / deň) sa môže vylúčiť
moč je normálna, oveľa viac - u detí a patológie.
Kreatín, kreatínfosfát a kreatinín
Kreatínfosfát - makroerg, akumulátor a nosičenergie v bunke.
Kreatínkináza
kreatín
+ ATP
~2%
ENERGIE
pH
H20
KREATÍN~
FOSFÁT
+ ADP
Kreatinín
Kreatinín je konečným produktom metabolizmu ("metabolická slepá ulička").Vylučuje sa obličkami močom.
Hladiny kreatinínu v plazme priamo súvisia
svalová hmota. Pretože referenčné hodnoty kreatinínu v krvi
závisí od veku a pohlavia.
Vekové skupiny
Krv z pupočnej šnúry
Od Jaffa
Enzymatické
53-106 umol/l
Novorodenci 1-4 dni
27-88
Deti do 1 roka
18-35
4-29
deti
27-62
2-5 rokov 4-40
6-9 rokov 18-46
Tínedžeri
44-88
19-52
Dospelí 18-60 rokov
m 80-115, š 53-97
Dospelí vo veku 60-90 rokov
m 71-115, š 53-106
Dospelí > 90 rokov
m 88-150, š 53-115
m 55-96, š 40-66
Metódy na štúdium kreatinínu
2-bodová kinetická metóda reakciou s kyselinou pikrovou(Jaffeho metóda).
Kreatinín + pikrát (pH=12,0) ---> oranžový produkt
Požiadavky na vzorku: Sérum alebo heparínová plazma (fluorid a
heparinát amónny sú nevhodné). Stabilné pri chladení
počas dňa, na dlhodobé skladovanie – zamraziť.
Príprava pacienta.
Odstráňte svalové napätie, cvičenie.
Strava by nemala obsahovať veľké množstvo mäsa.
Vyhnite sa alkoholu, veľkým dávkam kyseliny askorbovej,
ak je to možné - nefrotoxické lieky.
Analýza by sa mala vykonať pred rádiokontrastnými vyšetreniami.
Rušenie
Biologické: zvýšenie - nefrotoxické lieky.Analytické: zvýšenie - kyselina acetoctová, acetón,
kyselina askorbová, cefalosporíny, flucytozín, lidokaín,
ibuprofén, levodopa, metyldopa, nitrofurány, pyruvát, močové cesty
kyselina.
Falošné zvýšenie - glukóza, fruktóza, ketolátok, histidín, asparagín, močovina, kyselina močová, indol.
Znížené - n-acetylcysteín, bilirubín, dipyrón, hemoglobín,
lipémia.
Kolísanie ukazovateľa počas dňa môže dosiahnuť 100%
(maximálne - večer).
Klinická a diagnostická hodnota
ZVÝŠENÁ KONCENTRÁCIA KREATINÍNU:Zvýšené vzdelanie/vstupné
Akromegália a gigantizmus (veľká svalová hmota).
Nadmerná konzumácia mäsových potravín.
Znížený výber
Zlyhanie obličiek (akútne a chronické, akejkoľvek etiológie)
- porušenie perfúzie, ochorenie obličiek, obštrukcia močových ciest).
nefrotoxické látky - zlúčeniny ortuti, sulfónamidy,
tiazidy, aminoglykozidy, tetracyklíny, barbituráty, salicyláty,
androgény…
Mechanické, prevádzkové a iné rozsiahle poškodenia
svaly, predĺžený crush syndróm.
Choroba z ožiarenia, hypertyreóza.
Klinická a diagnostická hodnota
ZNÍŽENÁ KONCENTRÁCIA KREATINÍNUHladovanie.
Užívanie glukokortikoidov.
Slabosť spôsobená vekom alebo zníženou svalovou hmotou
omši.
Tehotenstvo (najmä prvý a druhý trimester).
Kreatinín nie je citlivý
ochorenie obličiek v počiatočnom štádiu.
Pri použití enzymatických metód treba byť opatrný
oddeľte sérum od buniek, aby ste zabránili jeho tvorbe
amoniaku vo vzorke a nadhodnotenie výsledkov.
Testy
1. Najväčší počet amoniak sa vylučuje z tela ako súčasť dusíkatej zložky moču:
Kreatinín. amónne soli. Indicana. Močovina . kyselina močová. Urobilinogén.
2. Pri výmene aminokyselín metionínu a serínu, ako zdroja jednouhlíkových radikálov v biosyntetických procesoch, sa vitamíny aktívne podieľajú ako koenzýmy:
Vitamín C. Vitamín D. Vitamín B12. Vitamín K. Tiamín. Kyselina listová. Vitamín RR. Riboflavín.
3. Ketogénne aminokyseliny zahŕňajú:
Serin. Valin. Leucín. metionín. izoleucín . histidín. Lyzín, tyrozín.
4. V dôsledku porušenia metabolizmu aminokyselín sa vyvíjajú choroby:
Fruktosémia. Dna. Alkaptonúria. Myxedém. albinizmus, fenylketonúria. Rachitída.
5. K fenylpyruvickej oligofrénii (fenylketonúrii) dochádza k porušeniu metabolizmu aminokyselín:
tyrozín. lyzín. fenylalanín. histidín. arginín.
6. Dôvodom rozvoja alkaptonúrie je porušenie metabolizmu aminokyselín:
cysteín. tryptofán. tyrozín. metionín. histidín. arginín.
7. Výraz "glykogénne aminokyseliny" znamená:
Znížiť renálny prah pre glukózu a spôsobiť glukozúriu. Narúšajú schopnosť buniek absorbovať glukózu. Schopný premeniť sa na glukózu a glykogén. Z energetického hľadiska dokážu nahradiť glukózu. Môže inhibovať proces glukoneogenézy.
8. Amoniak sa neutralizuje v pečeni tým, že je zahrnutý do syntézy močoviny v pečeni, látky sa priamo podieľajú:
Oxid uhličitý . lyzín. Ornitín.ATP. Kyselina glutámová. Aspartát, amoniak. Kyselina oxalooctová.
9. Na neutralizácii toxického amoniaku sa môže podieľať:
Kyselina acetoctová. Veveričky. Monosacharidy. Kyselina glutámová Kyselina alfa-ketoglutarová. Kyselina mliečna.
10. Čierna farba moču sa pozoruje pri ochorení:
Dna. fenylketonúria. Alkaptonúria . Žltačka
11. V prípade alkaptonúrie je enzým defektný:
Fenylalanín monooxygenáza. Dioxygenáza (oxidáza) kyseliny homogentisovej. Hydroláza kyseliny fumarylacetooctovej
12. Ktorý enzým je pri fenylketonúrii defektný?
Fenylalanín monooxygenáza. tyrozináza. Hydroláza kyseliny fumariaacetooctovej
13. Pri albinizme je metabolizmus tyrozínu narušený:
Oxidácia a dekarboxylácia. transaminácia
14. Pri tyrozinóze sú enzýmy defektné:
Hydroláza kyseliny fumarylacetooctovej. Tyrozíntransamináza
15. Minimálny podiel plnohodnotných bielkovín v strave dieťaťa z ich celkového príjmu by mal byť:
50%. 75%. 20%
Situačné úlohy
1. Mladá mamička informovala lekára o stmavnutí plienok počas ich sušenia. O čom dedičné ochorenie môžeš myslieť? Aké sú diétne odporúčania detského lekára?
2. 27. 36 hodín po narodení u chlapca zistili poruchu vedomia a dýchania. Pôrod je prirodzený, načas. rodičia - bratranci a sesternice a sestra. V krvnom sére bol zistený obsah amoniaku nad 1000 uM/l (norma 20-80), obsah močoviny bol 2,5 mmol/l (norma 2,5-4,5). Obsah kyseliny orotovej v moči je zvýšený. Dieťa zomrelo o 72 hodín neskôr.
V prospech čoho vrodené chyby výmena doložená laboratórnymi údajmi?
3. U 5-ročného dieťaťa po infekčná hepatitída obsah močoviny v krvi bol 1,9 mmol/l. O čom to svedčí túto analýzu? Aké sú odporúčania pediatra?
4. V prvých dňoch po pôrode má novorodenec vracanie, kŕče, v krvi bol zistený prudký nárast obsahu aminokyseliny ornitínu, veľmi nízka je koncentrácia močoviny. Akú chorobu má dieťa? Aké odporúčania možno použiť
5. Pacient s diabetes mellitus mal vysoký obsah močoviny v krvi. Počas obdobia zhoršenia celkového stavu sa však jeho koncentrácia v krvi z nejakého dôvodu znížila. Vysvetlite príčiny kolísania hladiny močoviny v krvi.
7. 1,5-mesačné dieťa má letargiu a letargiu. Vyšetrením bol zistený obsah fenylalanínu v krvi 35 mg/dl (normálne 1,4-1,9 mg/dl), obsah fenylpyruvátu v moči 150 mg/deň (normálne 5-8 mg/deň). Urobte záver o chorobe, jej príčine. Aké diétne odporúčania sú v tomto prípade potrebné?
8. 22-ročný pacient s arginínovou sukcinaturiou bol úspešne liečený ketoanalógmi aminokyselín fenylalanínu, valínu a leuínu na pozadí nízkobielkovinovej diéty. Koncentrácia amoniaku v plazme sa súčasne znížila z 90 na 30 µmol/l a výrazne sa znížilo vylučovanie arginín sukcinátu. Vysvetlite mechanizmus terapeutického účinku ketoanalógov aminokyselín.
9. Pri dedičnom ochorení familiárnej hyperamonémie dochádza k trvalému zvýšeniu obsahu amoniaku v krvi a úplnej absencii citrulínu. Hlavné klinické prejavy spojené s poškodením CNS. Aká reakcia je pri tejto chorobe blokovaná? Ako sa zmení denné vylučovanie močoviny?
10. V moči pacienta bolo zistené významné množstvo kyseliny homogentisovej. Aký dedičný enzymatický defekt možno predpokladať? Napíšte reakciu zablokovanú na tohto pacienta. Aké sú diétne odporúčania pre tohto pacienta?
Aké sú poruchy trávenia bielkovín v gastrointestinálnom trakte? Aké dodatočné testy sú potrebné?
11. Množstvo bielkovín v strave detí vo veku 3 a 13 rokov odporúča lekár v dávke 2,3 g/kg telesnej hmotnosti.
12. Dieťa bolo prijaté na detskú kliniku a potrebuje analýzu. tráviace šťavy. Zavedenie sondy je náročné. Ako vykonať štúdiu sekrečnej funkcie žalúdka?
23. Detský lekár predpísal pepsín dieťaťu s ochorením žalúdka. Aký liek je potrebný navyše? prečo?
13. S jedlom sa do tela tínedžera dostáva 80 g bielkovín denne. Počas tejto doby sa močom vylúčilo 16 g dusíka. Aká je dusíková bilancia u dieťaťa? O čom svedčí?
14. S močom fyzicky silného študenta strednej školy,
15 g dusíka. Mám zmeniť množstvo bielkovín v jeho strave?
15. Dieťa bolo prijaté na chirurgické oddelenie s bolesťami brucha. Laboratórne vyšetrenie odhalilo prudké zvýšenie indikanu v moči. Aký je možný dôvod tohto porušenia?
16. Matka dieťaťa trpiaceho nízkou kyslosťou žalúdočnej šťavy namiesto kyseliny chlorovodíkovej, ktorá mu bola predpísaná, začala používať roztok kyseliny citrónovej.
Je takáto náhrada možná? Vysvetlite prípustnosť alebo neprípustnosť tohto nahradenia.
Otázky na záverečnú hodinu na tému "Metabolizmus bielkovín a aminokyselín"
1. Vlastnosti metabolizmu bielkovín a aminokyselín. dusíková bilancia. faktor opotrebovania tela. Minimálne množstvo bielkovín. Kritériá nutričná hodnota bielkoviny. Proteínová diéta u malých detí. Kwashiorkor.
2. Trávenie bielkovín. Proteinázy gastrointestinálneho traktu a ich proenzýmy. Substrátová špecifickosť proteináz. Endo- a exopeptidázy. absorpcia aminokyselín. Vekové charakteristiky procesov trávenia a vstrebávania bielkovín .
3. Hnitie bielkovín v hrubom čreve. Produkty rozpadu a mechanizmy ich neutralizácie v pečeni. Vlastnosti priebehu hnilobných procesov v hrubom čreve dojčiat .
4. Dynamický stav bielkovín v tele. katepsíny. Autolýza tkanív a úloha poškodenia lyzozómov v tomto procese. Zdroje a hlavné spôsoby spotreby aminokyselín. Oxidačná deaminácia aminokyselín. Oxidáza aminokyselín, glutamátdehydrogenáza. Iné typy deaminácie aminokyselín.
5. Transaminácia. Aminotransferázy a ich koenzýmy. Biologický význam transaminačných reakcií. Špeciálnu úlohu v tomto procese má a-ketoglutarát. Nepriama deaminácia aminokyselín. Klinický význam stanovenia aktivity transamináz v krvnom sére.
6. Dekarboxylácia aminokyselín a ich derivátov. Najdôležitejšie biogénne amíny a ich biologická úloha. Rozpad biogénnych amínov v tkanivách.
7. Konečné produkty metabolizmu dusíka: amónne soli a močovina. Hlavné zdroje amoniaku v tele. Neutralizácia amoniaku. Biosyntéza močoviny (ornitínový cyklus). Vzťah medzi ornitínovým cyklom a Krebsovým cyklom. Pôvod atómov dusíka močoviny. Denné vylučovanie močoviny. Porušenie syntézy a vylučovania močoviny. Hyperamonémia. Vekové charakteristiky vylučovania dusíka konečných produktov z tela dieťaťa mladšieho ako 1 rok.
8. Neutralizácia amoniaku v tkanivách: redukčná aminácia a-ketokyselín, amidácia proteínov, syntéza glutamínu. Špeciálna úloha glutamínu v tele. Glutamináza obličiek. Adaptačná zmena aktivity glutaminázy obličiek pri acidóze.
9. Vlastnosti metabolizmu fenylalanínu a tyrozínu. Použitie tyrozínu na syntézu katecholamínov, tyroxínu a melanínov. Rozklad tyrozínu na kyselinu fumarovú a acetooctovú. dedičné poruchy výmena fenylalanínu a tyrozínu: fenylketonúria, alkaptonúria, albinizmus.
10. Vlastnosti metabolizmu serínu, glycínu, cysteínu, metionínu. Hodnota kyseliny tetrahydrolistovej a vitamínu B 12 v metabolizme jednouhlíkových radikálov. Nedostatok kyseliny listovej a vitamínu B12. Mechanizmus bakteriostatického účinku sulfátových liečiv.
11. Vzťah medzi metabolizmom aminokyselín a metabolizmom sacharidov a tukov. Glykogénne a ketogénne aminokyseliny. Nahraditeľné a nenahraditeľné aminokyseliny. Biosyntéza aminokyselín zo sacharidov.
ŠTRUKTÚRA A METABOLIZMUS NUKLEOVÝCH KYSELÍN
1. Zloženie RNA zahŕňa dusíkaté zásady:
adenín. guanín. uracil . Timin. Cytozín.
2. Jednotlivé nukleotidy v polynukleotidovom reťazci sú spojené väzbami:
Peptid. Fosfodiester. Disulfid. Vodík.
3. Enzýmy sa podieľajú na trávení nukleových kyselín - základných zložiek potravinových nukleoproteínov:
pepsín. Ribonukleáza. trypsín. Fosfolipázy. Deoxyribonukleáza. Amylase. Nukleotidáza. Fosfatázy.
4. Nukleové kyseliny majú najmenšiu molekulovú hmotnosť:
DNA. rRNA. tRNA. mRNA.
5. Konečným produktom rozkladu purínových dusíkatých zásad v ľudskom organizme je:
6. Hodnota denného vylučovania kyseliny močovej močom u dospelého zdravého človeka je:
0,01-0,05 g 0,06-0,15 g 0,35-1,5 g 2,5-5,0 g
7. Konečným produktom rozkladu pyrimidínových dusíkatých zásad v ľudskom tele je:
Močovina. Kyselina močová. amónne soli. Kreatinín
8. V rozpore s výmenou purínových dusíkatých zásad? Može sa stať patologických stavov:
Dna. Basedowova choroba. Choroba urolitiázy. Lesh-Nihanova choroba. Hyperamonémia.
9. stavebný materiál v matricovej syntéze nukleových kyselín sú látky:
Nukleozidové monofosfáty. Nukleozidové difosfáty. Nukleozidtrifosfáty. cyklické nukleotidy.
1. Proces biosyntézy RNA sa nazýva:
11. Biosyntéza bielkovín, uskutočňovaná za účasti polyzómov a tRNA, sa nazýva:
Prepis. Vysielanie. Replikácia. Oprava. Rekombinácia.
12. Hlavný spôsob reprodukcie genetickej informácie sa nazýva:
Prepis. Vysielanie. Replikácia. Oprava. Rekombinácia.
13 Transformácia pro-RNA na „zrelé“ formy sa nazýva:
Rekombinácia. Spracovanie. Replikácia. Vysielanie. Ukončenie.
14. Spracovanie a -RNA, t.j. jej zrelosť je:
Odstránenie intrónov. Odstránenie exónov. Špecifická modifikácia (metylácia, deaminácia atď.).
15 "Nezmysel - kodóny" (nezmyselné kodóny) v štruktúre mRNA sú signálom:
Signál na spustenie syntézy bielkovín. Mutantne zmenený kodón. Signál na ukončenie syntézy bielkovín. Signál na pripojenie protetických skupín k syntetizovanému proteínu.
16. Výraz „degenerácia“ genetického kódu znamená:
Schopnosť aminokyseliny byť kódovaná viac ako jedným kodónom. Schopnosť kodónu kódovať viacero aminokyselín. Obsah štyroch nukleotidov v kodóne. Obsah dvoch nukleotidov v kodóne.
17. Chargaffove pravidlá charakterizujúce znaky dvojzávitnicovej štruktúry DNA zahŕňajú:
A = T. G = C. A = C. G = T. A + G = C + T. A + T = G + C.
17. Na syntézu pyrimidínových báz de novo sa používajú tieto látky:
Oxid uhličitý. Glutamát. Glutamín. aspartát. alanín.
19. Na tvorbu purínového cyklu pri syntéze purínových nukleotidov sa používajú tieto látky:
Oxid uhličitý. aspartát. alanín. Glycocol. Glutamín. Deriváty tetrahydrofolátu.
20. Špecifickosť interakcie aminokyselín s t-RNA je spôsobená:
Zloženie antikodónu. Znak štrukturálnej organizácie tRNA. Špecifickosť aminoacyl-tRNA syntetáz. Štruktúra aminokyseliny.
21. Na syntézu pyrimidínových nukleotidov sa používajú:
CO2. Glutamín. aspartát. alanín
22. Prekurzorom syntézy purínových nukleotidov sú:
Kyselina inozínová. Kyselina orotová. Kyselina močová
23 Orotacidúria sa vyvinie, keď „blok“ enzýmu:
Karbamoyl aspartát transferáza. Orotát fosforibozyltransferáza
Xantín oxidáza.
24. Prvým krokom v syntéze pyrimidínového kruhu je:
Karbamoylfosfát. Ribóza-5-fosfát. Kyselina orotová. aspartát
25. Nukleotid – prekurzor pri syntéze pyrimidínových nukleotidov je:
Inozínmonofosfát. Orotát monofosfát. kyselina xantylová. Kyselina orotová
26. Kľúčovými enzýmami pri syntéze pyrimidínových nukleotidov sú:
27. Kľúčové enzýmy v syntéze pri syntéze purínových nukleotidov sú:
Karbamoylfosfosyntetáza. Karbamoyl aspartát transferáza. Fosforibozyl amidotransferáza
28. Pri imunodeficienciách je aktivita enzýmov znížená:
Adenozíndeamináza. Xantín oxidáza. Purínová nukleozidová fosforyláza
29. Pri Lesh-Nyhanovom syndróme je aktivita enzýmu znížená:
Xantín oxidáza. Adenín fosforibozyltransferáza. Hypoxantín-guanín fosforibozyltransferáza
30. Enzýmová aktivita je znížená pri orotacidúrii:
Orotát fosforibozyltransferáza. Dihydroorotát dehydrogenáza. Karbamoyl aspartát transferáza
31. Proces premeny pro-RNA na zrelé formy sa nazýva:
Rekombinácia. Spracovanie. Vysielanie. Ukončenie. replikácie
32. Keď dôjde k spájaniu:
Vystrihovanie kópií intrónov. Excízia kópií exónov. Spojenie informatívnych oblastí RNA
33. Na prepis potrebujete:
DNA. Primer. RNA polymeráza. proteínové faktory. Nukleotid trifosfáty. topoizomeráza
34. Enzýmy sa podieľajú na syntéze RNA:
RNA polymeráza. DNA polymeráza. Topoizomerázy. Primas
35. "Exóny" pro-RNA sa nazývajú:
nekódujúce regióny. doplnkové proteíny. miesto terminálu. kódovacie oblasti. Začiatok stránky
36. Enzýmy sa podieľajú na oprave DNA:
DNA ligázy. DNA polymerázy.) DNA reštrikčné enzýmy. Primas
37. Replikácia vyžaduje:
DNA. Primer. I-RNA. proteínové faktory. Nukleotid trifosfáty.
T opoizomeráza
38. Enzýmy sa podieľajú na syntéze DNA:
RNA polymeráza. DNA polymeráza. Peptidyltransferázy. tTopoizomerázy. Primas
39. Na regulácii syntézy bielkovín sa podieľajú:
Regulačný gén. Exon. Operátorský gén. Represor. Intron. Štrukturálny gén
40. Pri posttranslačnej modifikácii proteínov sú možné nasledovné:
čiastočná proteolýza. Glykozylácia. modifikácia aminokyselín. Upevnenie protetickej skupiny
41. Proces pohybu mRNA pozdĺž ribozómu sa nazýva:
Translokácia. Vysielanie. Ukončenie
42. Enzým sa podieľa na tvorbe peptidovej väzby počas biosyntézy bielkovín:
Peptidyltransferáza. topoizomeráza. helicase
43. Signál pre začiatok a koniec syntézy polypeptidového reťazca je:
určité kodóny mRNA. určité enzýmy. Určité aminokyseliny
44. Denné vylučovanie močoviny u dospelého človeka je:
1,0-2,0 g 20,-30,0 g 2,0-8,0 g 35,0-50,0 g 8,0-20,0 g
0,1-0,3 mM/l. 0,17-0,41 mM/l. 0,05-0,1 mM/l
46. Podiel dusíka kyseliny močovej v moči detí je:
1-3%. 3-8,5 %. 0,5-1,0 %.
47. Podiel močovinového dusíka v moči novorodencov je:
30% . 75% . 50%.
Situačné úlohy
1. Pacient sa sťažuje na bolesti kĺbov. Obsah kyseliny močovej v krvi je 0,26 mmol / l. Množstvo sialových kyselín - 4,5 mmol / l
(norma 2,0-2,6 mmol / l). Aké ochorenie možno vylúčiť?
2. Dieťa má genetický defekt enzýmu hypoxantín-guanín fosforibozyltransferáza. K akým dôsledkom to môže viesť?
3. Pacient sa sťažuje na bolesti kĺbov. Obsah kyseliny močovej v krvi je 0,56 mmol / l. Množstvo sialových kyselín je 2,5 mmol/l (norma je 2,0-2,6 mmol/l). Aké ochorenie je najpravdepodobnejšie? Aká diéta sa zobrazuje?
4. V dôsledku génovej mutácie sa mení poradie striedania nukleotidov v kodóne. K čomu to môže viesť?
5. U dieťaťa trpiaceho hypovitaminózou je znížená výmena nukleových kyselín. Vysvetlite dôvody porušení. Ktoré vitamíny sú zobrazené ako prvé?
6. Pri diabetes mellitus sa rýchlosť syntézy nukleových kyselín výrazne znižuje. Popíšte možné príčiny tohto porušenia.
7. V dôsledku génovej mutácie sa mení poradie striedania nukleotidov v kodóne. K čomu to môže viesť?
8. Nádorové bunky sa vyznačujú zrýchleným delením a rastom buniek. Ako sa tomu dá predísť ovplyvňovaním syntézy dusíkatých zásad?
Otázky na záverečnú hodinu na tému „Výmena nukleoproteínov“
1. Nukleové kyseliny ako polymérne zlúčeniny. Zloženie a štruktúra nukleotidov, ich funkcie v organizme. Biologický význam nukleových kyselín. Úrovne štrukturálnej organizácie. Druhová špecifickosť primárnej štruktúry.
2. Hlavné typy nukleových kyselín v tkanivách. Ich všeobecné charakteristiky. Zvláštnosti chemické zloženie, štruktúra a vlastnosti molekúl DNA. Komplementárnosť dusíkatých zásad. Denaturácia a regenerácia DNA. Hybridizácia DNA"DNA a DNA"RNA.
3. Rozpad v tkanivách pyrimidínových a purínových nukleotidov. konečné produkty rozkladu. Vlastnosti vylučovania kyseliny močovej z tela. Hyperurikémia. Dna.
4. Biosyntéza pyrimidínových nukleotidov. Alosterické mechanizmy regulácie.
5. Biosyntéza purínových nukleotidov. Pôvod častí purínového jadra. Počiatočné štádiá biosyntéza. Kyselina inozínová ako prekurzor kyseliny adenylovej a guanilovej. Alosterické mechanizmy regulácie biosyntézy.
6. biosyntéza DNA. Replikácia a oprava poškodenia. DNA biosyntetické enzýmy. Matrix. Korešpondencia primárnej štruktúry reakčného produktu s primárnou štruktúrou matrice. Semená (primer). Maticová úloha RNA. Revertase.
7. biosyntéza RNA. RNA polymeráza. Transkripcia je prenos informácií z DNA do RNA. Tvorba primárneho prepisu, jeho dozrievanie (spracovanie).
8. Biosyntéza bielkovín. Maticová (informačná) RNA. Základný postulát molekulárnej biológie: DNA®iRNA®proteín. Zhoda nukleotidovej sekvencie génu s aminokyselinovou sekvenciou proteínu (kolinearita). Problém prekladu (prekladu) štvormiestneho nukleotidového záznamu informácie do dvadsaťmiestneho aminokyselinového záznamu. Charakteristika nukleotidového kódu.
9. Transportná RNA (tRNA), štruktúrne a funkčné vlastnosti. Izoakceptorové formy tRNA. Biosyntéza aminoacyl-tRNA. Význam vysokej substrátovej špecifickosti aminoacyl-tRNA syntetáz.
10. Biologické systémy biosyntézy bielkovín. Štruktúra ribozómu. Sekvencia dejov v biosyntéze polypeptidového reťazca. Iniciácia, predĺženie, ukončenie. Regulácia biosyntézy bielkovín. Inhibítory biosyntézy matrice: lieky, vírusové a bakteriálne toxíny. Posttranslačná zmena v polypeptidovom reťazci.
 Metabolizmus dusíka - súbor chemických premien látok obsahujúcich dusík v tele. A. o. zahŕňa výmenu jednoduchých a zložitých bielkovín, nukleových kyselín, produktov ich rozpadu (peptidy, aminokyseliny a nukleotidy), tukom podobných látok s obsahom dusíka (lipidov), aminocukrov, hormónov, vitamínov atď. Pre normálny priebeh vit. procesov, treba telu poskytnúť potrebné množstvo stráviteľný dusík. hlavný neoddeliteľnou súčasťou a hlavným zdrojom dusíka v ľudskej potrave sú bielkovinové látky
Metabolizmus dusíka - súbor chemických premien látok obsahujúcich dusík v tele. A. o. zahŕňa výmenu jednoduchých a zložitých bielkovín, nukleových kyselín, produktov ich rozpadu (peptidy, aminokyseliny a nukleotidy), tukom podobných látok s obsahom dusíka (lipidov), aminocukrov, hormónov, vitamínov atď. Pre normálny priebeh vit. procesov, treba telu poskytnúť potrebné množstvo stráviteľný dusík. hlavný neoddeliteľnou súčasťou a hlavným zdrojom dusíka v ľudskej potrave sú bielkovinové látky

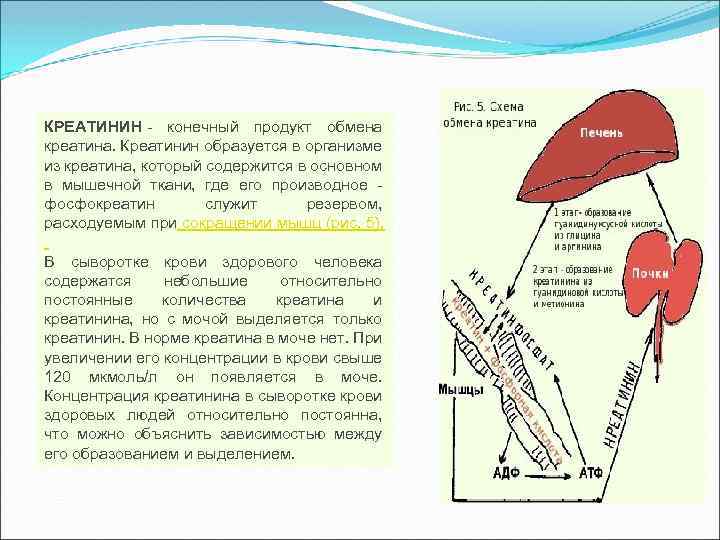 Kreatinín je konečným produktom metabolizmu kreatínu. Kreatinín sa v tele tvorí z kreatínu, ktorý sa nachádza najmä v svalové tkanivo, kde jeho derivát, fosfokreatín, slúži ako rezerva spotrebovaná pri svalovej kontrakcii (obr. 5). Krvné sérum zdravého človeka obsahuje malé, relatívne konštantné množstvá kreatínu a kreatinínu, ale len kreatinín sa vylučuje močom. Normálne nie je v moči žiadny kreatín. Pri zvýšení jeho koncentrácie v krvi nad 120 µmol / l sa objavuje v moči. Koncentrácia kreatinínu v sére zdravých ľudí relatívne konštantný, čo možno vysvetliť vzťahom medzi jeho tvorbou a uvoľňovaním.
Kreatinín je konečným produktom metabolizmu kreatínu. Kreatinín sa v tele tvorí z kreatínu, ktorý sa nachádza najmä v svalové tkanivo, kde jeho derivát, fosfokreatín, slúži ako rezerva spotrebovaná pri svalovej kontrakcii (obr. 5). Krvné sérum zdravého človeka obsahuje malé, relatívne konštantné množstvá kreatínu a kreatinínu, ale len kreatinín sa vylučuje močom. Normálne nie je v moči žiadny kreatín. Pri zvýšení jeho koncentrácie v krvi nad 120 µmol / l sa objavuje v moči. Koncentrácia kreatinínu v sére zdravých ľudí relatívne konštantný, čo možno vysvetliť vzťahom medzi jeho tvorbou a uvoľňovaním.
 Močovina je konečným produktom rozkladu bielkovín v tele, ktorý pri vylučovaní obličkami odstraňuje zvyšky „nepotrebného“ dusíka. Vzniká v pečeni. Vylučuje sa spolu s močom a čiastočne s potom (čo dodáva potu špecifický zápach). Močovina pomáha telu zadržiavať vodu a niektoré stopové prvky. To sa deje prostredníctvom reabsorpcie obličkami do krvného obehu. Zakaždým, keď dôjde k procesu absorpcie, močovina „stiahne“ molekuly vody späť do krvi a užitočné minerály. Jeho nadmerný obsah však poškodzuje orgány a tkanivá.
Močovina je konečným produktom rozkladu bielkovín v tele, ktorý pri vylučovaní obličkami odstraňuje zvyšky „nepotrebného“ dusíka. Vzniká v pečeni. Vylučuje sa spolu s močom a čiastočne s potom (čo dodáva potu špecifický zápach). Močovina pomáha telu zadržiavať vodu a niektoré stopové prvky. To sa deje prostredníctvom reabsorpcie obličkami do krvného obehu. Zakaždým, keď dôjde k procesu absorpcie, močovina „stiahne“ molekuly vody späť do krvi a užitočné minerály. Jeho nadmerný obsah však poškodzuje orgány a tkanivá.
 Kyselina močová Konečný produkt metabolizmu purínových báz, ktoré tvoria nukleotidy. V dôsledku vylučovania kyseliny močovej sa z tela odstraňuje nadbytočný dusík. V plazme sa kyselina močová nachádza prevažne vo forme sodnej soli. Koncentrácia kyseliny močovej v krvi je spôsobená rovnováhou procesov syntézy kyseliny močovej a jej vylučovania obličkami.
Kyselina močová Konečný produkt metabolizmu purínových báz, ktoré tvoria nukleotidy. V dôsledku vylučovania kyseliny močovej sa z tela odstraňuje nadbytočný dusík. V plazme sa kyselina močová nachádza prevažne vo forme sodnej soli. Koncentrácia kyseliny močovej v krvi je spôsobená rovnováhou procesov syntézy kyseliny močovej a jej vylučovania obličkami.
 SPÔSOBY TVORBY AMONIAKU kyselina glutámová - kyselina ketoglutárová + NH 3 - aminokyselina - ketokyselina + NH 3 cysteínpyruvát + NH 3 histidín kyselina urokaová + NH 3 glycín kyselina glyoxálová + NH 3 glukózamín-6-fosfát glukóza-6 - fosfát + NH 3 glutamín kyselina glutámová + NH 3 O ║ NH 2 -C O-P + CO 2 + NH 3 karbomoyl fosfát glutamín ATP ADP močovina pyrimidíny kyselina listová puríny glukozamín
SPÔSOBY TVORBY AMONIAKU kyselina glutámová - kyselina ketoglutárová + NH 3 - aminokyselina - ketokyselina + NH 3 cysteínpyruvát + NH 3 histidín kyselina urokaová + NH 3 glycín kyselina glyoxálová + NH 3 glukózamín-6-fosfát glukóza-6 - fosfát + NH 3 glutamín kyselina glutámová + NH 3 O ║ NH 2 -C O-P + CO 2 + NH 3 karbomoyl fosfát glutamín ATP ADP močovina pyrimidíny kyselina listová puríny glukozamín
Src="http://present5.com/presentation/229073585_437016682/image-8.jpg" alt="(!LANG:PRÍČINY INTOXIKÁCIE AMONIAKOM HYPERAMONIÉMIA (>25 -40 µmol/l) Porušenie funkcie tvorby močoviny pečeň Porušenie funkcie vylučovania dusíka"> ПРИЧИНЫ АММИАЧНОЙ ИНТОКСИКАЦИИ ГИПЕРАММОНИЙЕМИЯ (>25 -40 мкмоль/л) Нарушение мочевинообразующей функции печени Нарушение азотовыделительной функции почек Острая !} zlyhanie obličiek Vírusová hepatitída Chronické zlyhanie obličiek Toxická hepatitída Cirhóza pečene Portakaválne anastomózy na pozadí potravy bohatej na bielkoviny Vrodená hyperamonémia
 Zvyškový dusík a jeho zložky Indikátor Obsah v krvnom sére v mg/100 ml SI jednotky Zvyškový dusík 20-40 mg/100 ml 7,06-14,1 mmol/l Močovina 20-40 mg/100 ml 3,3-6, 6 mmol/l Dusík amino kyseliny 2,0-4,3 mg/100 ml 1,43-3,07 mmol/l Kyselina močová 2-6,4 mg/100 ml 0,12-0,38 mmol/l muži 0 , 2-0,7 mg/100 ml 13-53 µmol/l ženy 0,4-0,9 mg /100 ml 27-71 µmol/l muži 1-2 mg/100 ml 0,088-0,177 mmol /l ženy 0,5-1,6 mg/100 ml 0,044-0,141 mmol/l Amoniak 0,03-0,06 mg/100 ml: Kreatín 28,4 Kreatinín: Iné neproteínové látky (polypeptidy, nukleotidy atď.) Xantoproteínová reakcia 20 jednotiek. Kreatín: celá krv 3-4 mg % 229-305 µmol/l plazma 1- 1,5 mg% 76,3- 114,5 µmol/l Dusík močoviny v krvi (močovina: 2, 14) 9-14 mg % 3, 18- 4,94 mmol/l
Zvyškový dusík a jeho zložky Indikátor Obsah v krvnom sére v mg/100 ml SI jednotky Zvyškový dusík 20-40 mg/100 ml 7,06-14,1 mmol/l Močovina 20-40 mg/100 ml 3,3-6, 6 mmol/l Dusík amino kyseliny 2,0-4,3 mg/100 ml 1,43-3,07 mmol/l Kyselina močová 2-6,4 mg/100 ml 0,12-0,38 mmol/l muži 0 , 2-0,7 mg/100 ml 13-53 µmol/l ženy 0,4-0,9 mg /100 ml 27-71 µmol/l muži 1-2 mg/100 ml 0,088-0,177 mmol /l ženy 0,5-1,6 mg/100 ml 0,044-0,141 mmol/l Amoniak 0,03-0,06 mg/100 ml: Kreatín 28,4 Kreatinín: Iné neproteínové látky (polypeptidy, nukleotidy atď.) Xantoproteínová reakcia 20 jednotiek. Kreatín: celá krv 3-4 mg % 229-305 µmol/l plazma 1- 1,5 mg% 76,3- 114,5 µmol/l Dusík močoviny v krvi (močovina: 2, 14) 9-14 mg % 3, 18- 4,94 mmol/l
 produkčná hyperazotémia Hepatocelulárna insuficiencia nemočovinových frakcií zvyškového dusíka (amónny, amónny dusík, biogénne amíny); ↓frakcia močoviny v zvyškovom dusíku Posilnený proteínový katabolizmus (hladovanie, prekrmovanie) nemočovinových frakcií zvyškového dusíka (amónium, amónny dusík, biogénne amíny); frakcia močoviny pri retencii zvyškového dusíka Akútne a chronické zlyhanie obličiek AKI: koncentrácia močoviny v plazme, ↓klírens močoviny, zvyškového dusíka a močovinového dusíka CRF: zvyškový dusík do 200-300 mg/100 ml, vy, peptidy ("stredné molekuly") v plazme
produkčná hyperazotémia Hepatocelulárna insuficiencia nemočovinových frakcií zvyškového dusíka (amónny, amónny dusík, biogénne amíny); ↓frakcia močoviny v zvyškovom dusíku Posilnený proteínový katabolizmus (hladovanie, prekrmovanie) nemočovinových frakcií zvyškového dusíka (amónium, amónny dusík, biogénne amíny); frakcia močoviny pri retencii zvyškového dusíka Akútne a chronické zlyhanie obličiek AKI: koncentrácia močoviny v plazme, ↓klírens močoviny, zvyškového dusíka a močovinového dusíka CRF: zvyškový dusík do 200-300 mg/100 ml, vy, peptidy ("stredné molekuly") v plazme
 ZDROJE A METÓDY NEUTRALIZÁCIE AMONIAKU V RÔZNYCH TKANIVÁCH Biogénne amíny Aminokyseliny Nukleotidy AMONIAK Syntéza Tvorba močoviny glutamín alanín glutamát amónne soli (25 g/deň) (0,5 g/deň) pečeň mozog svaly, črevá mozog obličky
ZDROJE A METÓDY NEUTRALIZÁCIE AMONIAKU V RÔZNYCH TKANIVÁCH Biogénne amíny Aminokyseliny Nukleotidy AMONIAK Syntéza Tvorba močoviny glutamín alanín glutamát amónne soli (25 g/deň) (0,5 g/deň) pečeň mozog svaly, črevá mozog obličky
 REAKCIE VÄZBY AMONIAKU V BUNKE 1. Reakcia redukčnej aminácie -ketoglutarátu na L-glutamát: NADPH 2 NADPH NH 3 + -ketoglutarová kyselina glutámová + H 2 O kyselina glutamátdehydrogenáza kyselina 2. Reakcia vzniku glutamínu z kyseliny glutámovej za účasti enzýmu glutamínsyntetázy. Reakcia prebieha v cytosóle buniek všetkých tkanív, ale vo väčšej miere v mozgu: COOH CONH 2 │ │ CH 2 ATP ADP + Fn CH 2 │ │ CH 2 2 CH │ │ HC-NH 2 glutamín syntetáza HC- NH 2 │ │ COOH COOH glutamín glutamín kys
REAKCIE VÄZBY AMONIAKU V BUNKE 1. Reakcia redukčnej aminácie -ketoglutarátu na L-glutamát: NADPH 2 NADPH NH 3 + -ketoglutarová kyselina glutámová + H 2 O kyselina glutamátdehydrogenáza kyselina 2. Reakcia vzniku glutamínu z kyseliny glutámovej za účasti enzýmu glutamínsyntetázy. Reakcia prebieha v cytosóle buniek všetkých tkanív, ale vo väčšej miere v mozgu: COOH CONH 2 │ │ CH 2 ATP ADP + Fn CH 2 │ │ CH 2 2 CH │ │ HC-NH 2 glutamín syntetáza HC- NH 2 │ │ COOH COOH glutamín glutamín kys
 Cesty výmeny dusíkatých aminokyselín a amoniaku TKANIVO KRV PEČEŇ Svaly, črevá Amino-KG Ala-KG NH 3 Ketok-you glutamát pyruvát pyruvát glutamát CO 2 AMP NH 3 glukóza karbomoyl fosfát IMF glutamát glutamín Amino-K G BRAIN a iné tkanivá 3 Ketok-you glutamát glutamín -KG OBLIČKA NH 3 glutamát NH 3 MOČ ornitínový cyklus NH 3 glutamát močovina amónne soli glutamát močovina
Cesty výmeny dusíkatých aminokyselín a amoniaku TKANIVO KRV PEČEŇ Svaly, črevá Amino-KG Ala-KG NH 3 Ketok-you glutamát pyruvát pyruvát glutamát CO 2 AMP NH 3 glukóza karbomoyl fosfát IMF glutamát glutamín Amino-K G BRAIN a iné tkanivá 3 Ketok-you glutamát glutamín -KG OBLIČKA NH 3 glutamát NH 3 MOČ ornitínový cyklus NH 3 glutamát močovina amónne soli glutamát močovina
 CYKLUS MOČOVINY A JEHO VZŤAH S TCA cytoplazmou NH 3 + CO 2 + H 2 O L-aspartát 2 ATP karbomoyl fosfát citrulín argininosukcinát oxaloacetát mitochondrie malát mitochondrie ornitín arginín fumarát močovina TCA
CYKLUS MOČOVINY A JEHO VZŤAH S TCA cytoplazmou NH 3 + CO 2 + H 2 O L-aspartát 2 ATP karbomoyl fosfát citrulín argininosukcinát oxaloacetát mitochondrie malát mitochondrie ornitín arginín fumarát močovina TCA